
L’insulina è un ormone proteico di 52 aminoacidi fondamentale per la regolazione dei livelli ematici di glucosio (glicemia) e secreto esclusivamente dalla β-cellula pancreatica. L’insulina umana è costituita da due catene polipeptidiche, A e B, formate da 21 e 30 aminoacidi, rispettivamente. Le due catene polipeptidiche sono unite da due ponti disolfuro tra i residui di cisteina in posizione 7 della catena A e B e i residui di cisteina in posizione 20 della catena A e in posizione 19 della catena B. Un ulteriore ponte disolfuro lega i residui di cisteina in posizione 6 e 11 della catena A e tale legame aminoacidico è ritenuto importante per la determinazione della struttura terziaria dell’insulina e per il suo legame al recettore insulinico (Fig.1). L’insulina ha un peso molecolare pari a 5.8 KDa ed è in grado di aggregarsi in dimeri, esameri e in strutture cristalline più complesse in presenza di ioni zinco e di un basso pH, come osservato nei granuli secretori.
L’insulina viene sintetizzata all’interno della β-cellula pancreatica sotto forma di pre-proinsulina e poi trasformata in proinsulina all’interno del reticolo endoplasmatico. Successivamente la proinsulina viene scissa nel complesso del Golgi in due componenti, il peptide C e l’insulina, immagazzinati nei granuli secretori. I granuli secretori presenti nella β-cellula sono di due tipi: immaturi, contenenti minime quantità di insulina ed elevate quantità di proinsulina, e maturi, presenti in quantità maggiore e contenenti l’ormone attivo. Sebbene la biosintesi dell’insulina sia controllata da molteplici fattori, il metabolismo del glucosio rappresenta il più importante stimolo fisiologico per la trascrizione del gene dell’insulina e per la traduzione del suo mRNA.
Anche il processo di secrezione insulinica viene regolato dal glucosio stesso. Questo processo prevede l’attivazione di una serie di eventi biochimici che possono essere suddivisi in eventi prossimali, rappresentati dall’ingresso del glucosio nella β-cellula e dal suo metabolismo, ed eventi distali rappresentati dalla generazione di segnali mitocondriali ed elettrici, responsabili a loro volta dell’esocitosi dei granuli contenenti l’insulina.
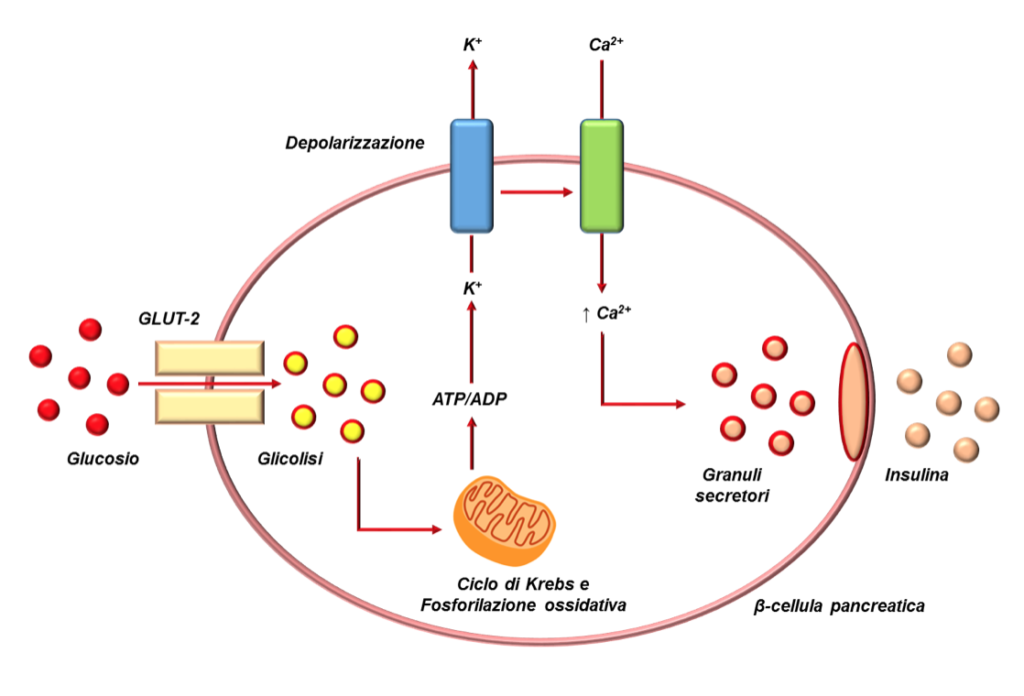
Il glucosio entra nella β-cellula pancreatica per mezzo del glucotrasportatore GLUT-2, la cui mobilizzazione verso la membrana plasmatica è insulino-indipendente e la cui affinità di legame per il glucosio è bassa; la molecola entra quindi nella glicolisi con la fosforilazione del glucosio in glucosio-6-fosfato da parte della glucochinasi e la produzione finale di due molecole di piruvato, due di NADH (Nicotinamide Adenina Dinucleotide Idrogenasi) e due di ATP (Adenosina Trifosfato) per ogni molecola di glucosio metabolizzata.
Grazie all’enzima piruvato-deidrogenasi, il piruvato viene convertito in acetil-CoA che entrando nel ciclo degli acidi tricarbossilici, detto anche ciclo di Krebs o ciclo dell’acido citrico, a livello mitocondriale dove subisce processi di ossidazione che generano CO2, GPT, FADH2 (Flavina Adenina Dinucleotide) e NADH. I coenzimi ridotti NADH e FADH2 trasportano elettroni fino alla catena respiratoria mitocondriale, dove vengono riossidati a NAD+ e FAD cedendo gli elettroni alla catena stessa, che sarà così in grado di rigenerare molecole di ATP a partire da ADP. L’ATP prodotto è responsabile della chiusura dei canali del potassio ATP-dipendenti (KATP) e della conseguente depolarizzazione della membrana della β-cellula, apertura dei canali del calcio (Ca2+) voltaggio-dipendenti ed esocitosi dei granuli di insulina (Fig.2). In questo caso si parla di secrezione di insulina indotta dal glucosio attraverso la via del canale KATP, Ca2+-dipendente. La secrezione insulinica può avvenire anche attraverso un canale KATP indipendente, Ca2+-dipendente quando l’aumento della concentrazione del Ca2+ intracellulare viene indotto da meccanismi differenti. Le due vie risultano sinergiche. Esiste anche una terza via indipendente sia da KATP sia dal Ca2+, ma dipendente dalla PKA (Protein-chinasi A) e dalla PKC (Protein-chinasi C).
Oltre al glucosio, alcuni aminoacidi e gli acidi grassi possono regolare la secrezione dell’insulina.
Gli aminoacidi presi singolarmente hanno un basso potere di indurre la secrezione dell’insulina; tuttavia, alcune combinazioni di aminoacidi, in concentrazioni fisiologiche o sovra-fisiologiche, possono incrementare la secrezione dell’insulina indotta dal glucosio da parte delle β-cellule pancreatiche. Un esempio è dato dalla combinazione glutamina-leucina. Gli acidi grassi possono incrementare la secrezione insulinica attraverso la sintesi di intermedi metabolici, quali l’acil-CoA a lunga catena e il diacilglicerolo (DAG), in grado di acilare le proteine coinvolte nella fusione dei granuli di insulina, tra cui la sinaptogamina, e di attivare PKC, rispettivamente. Anche alcuni ormoni, tra cui la melatonina, gli estrogeni, la leptina, il GH (Growth Hormone) e il GLP-1 (Glucagon Like Peptide-1) e alcuni neurotrasmettitori, quali l’acetilcolina e la noradrenalina, possono modulare la secrezione insulinica.
La secrezione dell’insulina consiste nella fusione dei granuli di insulina con la membrana plasmatica e nella successiva esocitosi del contenuto dei granuli. Si distinguono due tipi di granuli di insulina: i granuli immediatamente disponibili per il rilascio, che rappresentano l’1% di tutti i granuli e che contribuiscono al rilascio rapido dell’insulina indotto dal glucosio, e quelli di riserva. I granuli secretori traslocano fino alla membrana plasmatica mediante l’attivazione delle proteine contrattili actina e miosina che a loro volta interagiscono con i microtubuli con un meccanismo Ca2+-dipendente, e successivamente ha luogo il processo di fusione delle vescicole con la membrana plasmatica mediato dalle proteine SNARE (Solubile N-ethylmaleimide-sensitive factor Attachment protein Receptors) con liberazione dell’insulina nel torrente ematico. La secrezione insulinica si realizza in due fasi: la prima fase è transiente, si esaurisce in 10 minuti ed è legata alla frazione di granuli immediatamente disponibili per il rilascio, la seconda fase è più lenta e progressiva e comporta la produzione di nuova insulina. Inoltre, la secrezione di insulina avviene in maniera pulsatile: circa ogni 12-14 minuti si ha l’immissione in circolo di una piccola quantità di insulina al fine di modulare costantemente la glicemia.
L’emivita dell’insulina è molto breve (4-6 minuti). La clearance dell’insulina si verifica soprattutto a livello renale, ma anche a carico del fegato e di altri tessuti, tra cui quello muscolare. Il meccanismo maggiormente studiato attraverso cui l’insulina viene degradata parte dalla internalizzazione del complesso insulina-recettore insulinico attraverso un processo di invaginazione della membrana plasmatica e si realizza con la degradazione enzimatica nelle vescicole endocitiche stesse oppure nel citosol, nell’apparato del Golgi oppure nel lisosoma. L’insulina può essere captata anche per pinocitosi, soprattutto quando la sua concentrazione risulta essere alta.

- Giorgino R. Il Diabete Mellito. SEU Società Editrice Universo; 2011. https://www.medicalinformation.it/il-diabete-mellito-giorgino-riccardo-seu-societa-editrice-universo-editore.html
- Duckworth W, Bennett R, Hamel F. Insulin degradation: progress and potential. Endocr Rev. 1998;19(5):608-624. doi:10.1210/edrv.19.5.0349
- Fu Z, Gilbert E, Liu D. Regulation of insulin synthesis and secretion and pancreatic Beta-cell dysfunction in diabetes. Curr Diabetes Rev. 2013;9(1):25-53. https://www.ncbi.nlm.nih.gov/pubmed/22974359
